
Alternative Pathway
- The alternate pathway bypasses C1, C4, C2 components and starts at C3 step.
- It is generally activated by non-immunological means such as bacterial surfaces and their components (e.g. endotoxin, microbial polysaccharides).
- Some other factors such as inulin, zymosan, yeast walls, cobra venom are also responsible for activation of this pathway.
- However, the mechanism can also be triggered by IgA, IgE and IgG.
- This pathway does not depend on antibody action and bypasses early complement components (C1, C2 andC4) due to which it can be activated before the establishment of an immune response to the infecting bacteria.
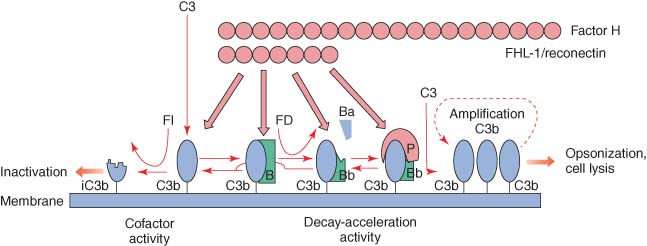
- Under normal situation (in the absence of infection), C3 is cleaved very slowly through reaction with water or trace amounts of proteolytic enzyme to give C3b and C3a.
- The C3b formed is susceptible to the nucleophilic attack by water and is rapidly inactivated to give iC3b.
- The C3a is not generated in sufficient amounts to lead to leukocyte activation and is rapidly inactivated.
- This normal low-level cleavage of the C3 molecule is termed ‘C3 tickover’ and it provides low levels of starting material i.e. C3b.
- This will be required for full activation of the alternative complement pathway in the case of microbial infection.
- In the presence of microbial membrane the C3b formed by C3 tick-over will be susceptibe to nucleophilic attack by hydroxyl or amine groups on the membrane surface.
- This leads to the co-valent attachment of C3b to the membrane.
- Once the C3b has attached the membrane, the initial activation of alternate pathway is mediated by factor B binding to C3b in the presence of Mg++ ions, which is then acted on by factor D.
- This factor splits factor B in the complex to yield the Bb active fragment that remains linked to C3b, i.e. formation of activation unit C3bBb.
- Inactive Ba is split from this complex, which leads to cleavage and activation of C3.
- C3 convertase (C3bBb) is capable of splitting more C3 to C3b, some of which becomes membrane bound.
- The complement cascade continues in a manner analogous to the classical pathway.
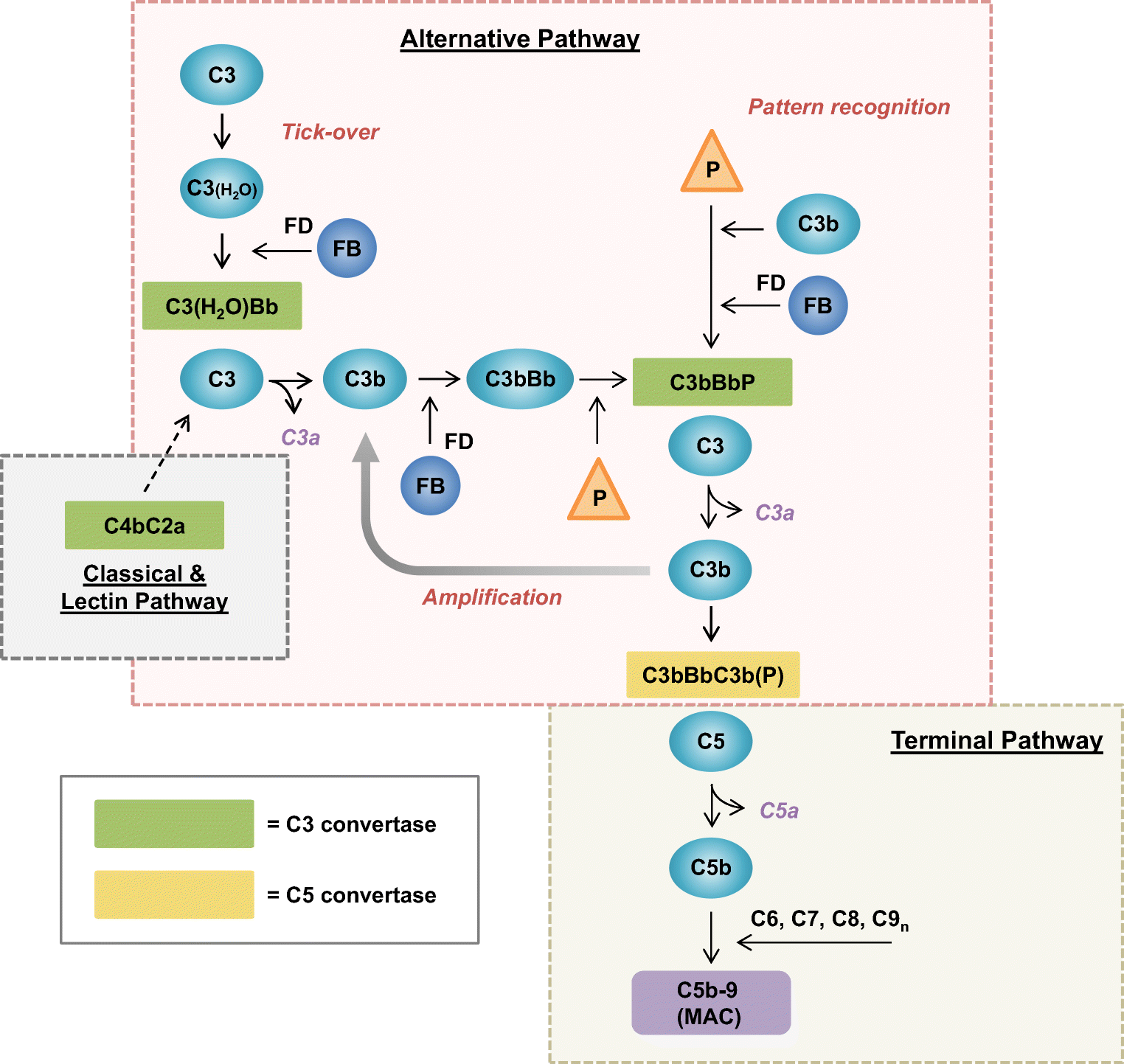
Image source: springerlink
Membrane attack complex
- The terminal stage of the complement pathway involves creation of the membrane attack complex, which is known as lytic unit.
- The C5 convetases are generated from C4b2a of the classical and C3bBb of the alternative pathway by addition of another C3b molecules.
- These membrane bound tri-molecular complexes selectively bind C5 and cleave it into a small C5a released in body fluids (anaphylatoxic and chemotactic) and a large membrane bound C5b.
- The formation of rest of the membrane attack complex is non-enzymatic.
- C6 binds to C5b and the joint complex C5b6 is released from the C5 convertase.
- C7 binds to the C5b6 complex forming a heat stable tri-molecular complex C5b67.
- The C5b67 is highly lipophilic and binds to membrane bilayer in the vicinity of initial activation site, where it lies as a high affinity receptor for C8.
- C5b678 binds and polymerizes C9 forming the membrane attack complex (MAC).
- As many as 12-16 C9 molecules can be polymerized by one C5b678 complex.
- A (C5b678)1(C9)n complex forms and drills a hole in the cell membrane, which leads to the hypotonic lysis of cells.
References:
i) https://www.svarlifescience.com/knowledge/focus-areas/complement-system-overview/alternative-pathway
ii) https://www.creative-biolabs.com/complement-therapeutics/alternative-pathway.htm